Le grain de pollen est généralement formé de seulement 2 cellules haploïdes : – la cellule végétative, responsable de l’allongement du tube pollinique, – la cellule spermatogène qui donnera 2 gamètes mâles ou spermatozoïdes, vecteurs de la fécondation. Le grain de pollen est entouré de 2 parois : – l’exine, épaisse, ornementée et lisse, – et l’intine, fine et doublant intérieurement l’exine. A certains endroits appelés apertures, l’exine est amincie. Chez le pollen des Monocotylédones ou des Dicotylédones basales (primitives), il n’existe normalement qu’une seule aperture, circulaire ou allongée en sillon. Parfois, cette aperture s’étire, s’amincie par endroits et forme alors plusieurs pores. Chez le pollen des Dicotylédones vraies (Eudicotylédones), il existe normalement 3 apertures. Cependant, ce nombre peut être multiplié. 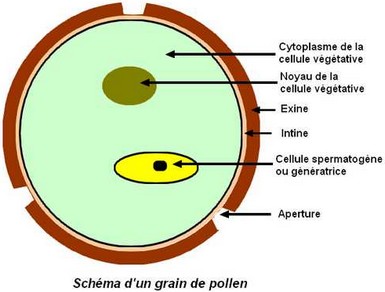
Archives par mot-clé : pollen
Pollinisation chez les Angiospermes
La pollinisation peut se faire selon différents modes : – l’autopollinisation. Le stigmate d’une fleur reçoit le pollen de la même plante. Ce mode est fréquent mais non obligatoire chez les Graminées cultivées. Il est par contre obligatoire chez les fleurs qui ne s’ouvrent pas (cléistogames) telles que la Violette. – la pollinisation croisée. Le stigmate d’une fleur reçoit le pollen d’une autre plante. La pollinisation croisée peut être favorisée : – par l’existence de fleurs mâles et de fleurs femelles sur des pieds différents, – par la présence d’organes reproducteurs n’ayant pas leur maturité en même temps : le pollen est libéré avant, alors que le stigmate est immature (protandrie) ou le stigmate est réceptif alors que les étamines sont encore jeunes (protogynie), – par l’existence de structures empêchant le pollen de se déposer sur le stigmate de la même fleur (rostellum des Orchis). Les agents de la pollinisation sont le vent (anémogamie) ou les insectes (entomogamie). Dans le premier cas, les fleurs sont en général à périanthe bien développé et coloré. Dans le second cas, le périanthe est absent ou réduit et peu coloré.
Germination du pollen
La germination du pollen commence par son gonflement par absorption de l’eau de la surface du stigmate. Les vacuoles turgescentes poussent l’intine et le cytoplasme vers une aperture du grain de pollen : c’est le début de la croissance du tube pollinique. Quand le tube pollinique s’agrandit le cytoplasme et les 2 noyaux se trouvent confinés à son extrémité, le noyau gamétogène étant légèrement en retrait du noyau végétatif. Les phénomènes d’élongation cellulaire se trouvent limités à l’extrémité du tube. Celui traverse le style sans jamais pénétrer dans les cellules ; les cellules du tissu de conduction agissent comme des rails pour le tube pollinique qui rentre au niveau du micropyle dans l’ovule.
La graine du Ginkgo (Ginkgo biloba)
Le Ginkgo est un arbre dioïque utilisé en arbre d’alignement car réputé résistant à la pollution. Les pieds femelles sont rarement utilisés car ils produisent des fruits qui, s’ils sont réputés avoir des vertus médicinales, sont nauséabonds. L’enveloppe charnue provient de la partie externe du tégument (sarcotesta) de l’ovule ou test et non pas d’un ovaire. Bien que les fruits du Ginkgo soient analogues aux fruits des Angiospermes, ils ne sont donc pas de même nature. Contrairement à ce qui se passe chez certaines Gymnospermes où les ovules fécondés sont protégés par des formations extra-ovulaires (pins, genevriers…), il n’y a pas de gymnospermie compensée et l’ovule reste nu : la gymnospermie de ginkgo est absolue.  Le tégument de l’ovule (test) du Ginkgo est divisé en trois parties : la couche externe (sarcotesta) est un parenchyme vascularisé, la couche médiane (sclérotesta) est dure car sclérifiée, la couche interne (endotesta) est fine et parcheminée. Avant la pollinisation, ce tégument laisse une ouverture, le micropyle, permettant au grain de pollen de s’insérer dans la chambre pollinique. Cette chambre produira un liquide mucilagineux dans lequel nageront les spermatozoïdes. Il y a fécondation aquatique comme chez certaines plantes primitives alors que le Ginkgo est une plante terrestre.
Le tégument de l’ovule (test) du Ginkgo est divisé en trois parties : la couche externe (sarcotesta) est un parenchyme vascularisé, la couche médiane (sclérotesta) est dure car sclérifiée, la couche interne (endotesta) est fine et parcheminée. Avant la pollinisation, ce tégument laisse une ouverture, le micropyle, permettant au grain de pollen de s’insérer dans la chambre pollinique. Cette chambre produira un liquide mucilagineux dans lequel nageront les spermatozoïdes. Il y a fécondation aquatique comme chez certaines plantes primitives alors que le Ginkgo est une plante terrestre.
Monocotylédones (Liliopsida)
A l’inverse des Dicotylédones, les Monocotylédones sont monophylétiques et forment un clade. Ils constituent la classe des Liliopsida. Les plantes sont monocotylédonées et présentent aussi les caractères suivants : – les formes herbacées sont majoritaires. Les géophytes à bulbes sont fréquentes ; – les bourgeons accessoires surnuméraires sont collatéraux, c’est-à-dire disposés sur un plan horizontal perpendiculaire à l’axe de la plante. Il y a une seule préfeuille adaxiale. Les bourgeons sériaux sont très rares ; – la ramification de la tige est rare (il ne faut pas la confondre avec celle de la hampe florale) ; – les feuilles sont normalement simples et linéaires, fréquemment disposées en rosette. La nervation est souvent parallèle, rarement pennée. Le bord du limbe est rarement denté. Les gaines et/ou les ligules sont fréquentes. Les feuilles sont rarement opposées ou verticillées (exception notable de Trillium) ; – la durée de vie de la racine principale est très faible et elle est très vite remplacée par des racines adventives ; – dans la tige, les faisceaux du phloème et du xylème sont très nombreux et sont disposés sur plusieurs cercles concentriques. Dans les racines les faisceaux sont ordinairement plus nombreux que chez les Dicotylédones ; – les tubes criblés du phloème sont toujours sans corpuscules lipo-protéiques ; – la croissance en épaisseur de la tige et de la racine est faible car les formations secondaires sont inexistantes sauf cas rares (Veratrum et certaines Monocotylédones arborescentes telles que les Yucca, les Dracaena, les Sansevieria, les Cordyline, les Aloe…). Dans ces derniers cas, la zone génératrice des formations secondaires n’est pas intrafasciculaire comme chez les Dicotylédones mais extra-fasciculaire ; – les fleurs sont généralement trimères et regroupées en des inflorescences qui peuvent être variées. La cyme unipare hélicoïde est, en particulier présente, chez les Zingiberaceae. Diverses inflorescences s’expriment aussi chez les Bromeliaceae ; – le pollen est uniaperturé ou d’un type dérivé, parfois pantoporé ; – la semence possède un seul plan de symétrie passant par le cotylédon et la radicule est coiffée d’un coléorhize.