L’ovule est constitué de l’extérieur vers l’intérieur: – de deux téguments, la primine, externe et vascularisée et la secondine, interne. Les téguments s’ouvrent au niveau du micropyle, – le nucelle, tissu diploïde, entouré des deux téguments. Si le nucelle est mince, l’ovule est tenuinucellé ; s’il est épais, il est crassinucellé, – le sac embryonnaire contenu dans le nucelle et constitué de 8 cellules haploïdes. 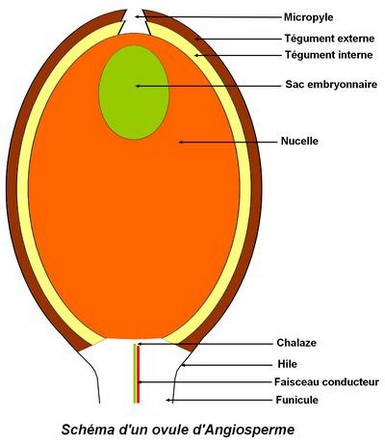
Archives par mot-clé : fécondation
Sac embryonnaire
Le sac embryonnaire est généralement constitué de 8 cellules haploïdes : – les 2 synergides et l’oosphère, près du micropyle, – les 3 cellules antipodiales, près de la chalaze, – les 2 noyaux polaires au centre du sac. Les noyaux polaires peuvent fusionner avant la fécondation. 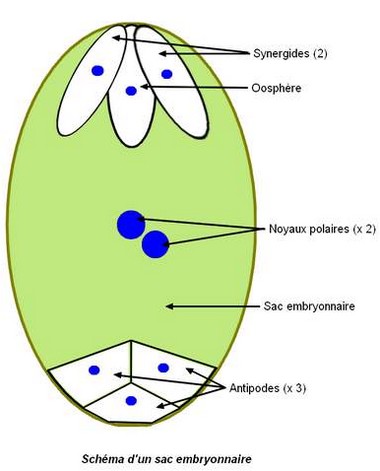
Pollen des Angiospermes
Le grain de pollen est généralement formé de seulement 2 cellules haploïdes : – la cellule végétative, responsable de l’allongement du tube pollinique, – la cellule spermatogène qui donnera 2 gamètes mâles ou spermatozoïdes, vecteurs de la fécondation. Le grain de pollen est entouré de 2 parois : – l’exine, épaisse, ornementée et lisse, – et l’intine, fine et doublant intérieurement l’exine. A certains endroits appelés apertures, l’exine est amincie. Chez le pollen des Monocotylédones ou des Dicotylédones basales (primitives), il n’existe normalement qu’une seule aperture, circulaire ou allongée en sillon. Parfois, cette aperture s’étire, s’amincie par endroits et forme alors plusieurs pores. Chez le pollen des Dicotylédones vraies (Eudicotylédones), il existe normalement 3 apertures. Cependant, ce nombre peut être multiplié. 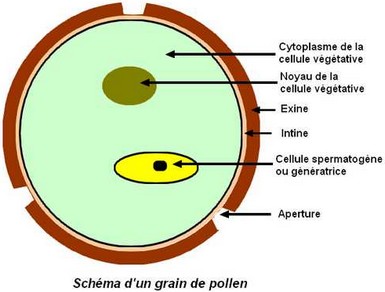
Zygote principal et albumen
Un noyau spermatique fusionne avec celui de l’oosphère. Le zygote principal obtenu donnera l’embryon. L’autre noyau spermatique fusionne avec les 2 noyaux centraux bien individualisés ou déjà fusionnés. Il en résulte un zygote accessoire ou albumen. L’albumen est triploïde mais, étant donné que les noyaux centraux ne sont pas toujours au nombre de deux ou diploïdes, son degré de ploïdie peut varier. L’albumen est un tissu de réserve qui termine sa formation avant celle du zygote principal. Quand ce dernier aura terminé sa propre formation, l’albumen aura généralement disparu (graine exalbuminée) ou subsistera (graine albuminée).
Pollinisation chez les Angiospermes
La pollinisation peut se faire selon différents modes : – l’autopollinisation. Le stigmate d’une fleur reçoit le pollen de la même plante. Ce mode est fréquent mais non obligatoire chez les Graminées cultivées. Il est par contre obligatoire chez les fleurs qui ne s’ouvrent pas (cléistogames) telles que la Violette. – la pollinisation croisée. Le stigmate d’une fleur reçoit le pollen d’une autre plante. La pollinisation croisée peut être favorisée : – par l’existence de fleurs mâles et de fleurs femelles sur des pieds différents, – par la présence d’organes reproducteurs n’ayant pas leur maturité en même temps : le pollen est libéré avant, alors que le stigmate est immature (protandrie) ou le stigmate est réceptif alors que les étamines sont encore jeunes (protogynie), – par l’existence de structures empêchant le pollen de se déposer sur le stigmate de la même fleur (rostellum des Orchis). Les agents de la pollinisation sont le vent (anémogamie) ou les insectes (entomogamie). Dans le premier cas, les fleurs sont en général à périanthe bien développé et coloré. Dans le second cas, le périanthe est absent ou réduit et peu coloré.