Pollination can be made according to various modes: – Self pollination. The stigma of a flower receives the pollen of the same plant. This mode is frequent, but not compulsory, in cultivated Grasses. It is on the other hand compulsory for flowers that do not open (cleistogamous ) such as the Violet. – Crossed pollination. The stigma of a flower receives the pollen of another plant. Cross pollination can be promoted: – By dioecism: male flowers and female flowers are on separate plants (dioecious species), – By dichogamy: male and female organs mature at different times. The pollen is before released while the stigma is immature (protandry) or the stigma is receptive while stamens are still young (protogyny), – By hercogamy: some structures prevent pollen from being transferred on stigma of the same flower (rostellum of the Orchis), – By heterostyly: in Primula, flowers with high style and stamens situated on the base of the corolla must be pollinated by flowers with short style and stamens situated on the top of the corolla, – By self sterility: flowers can’t be self pollinated because of dimorphism in pollen grains and stigma surfaces. Means of pollination are the wind (anemophily) or insects (entomophily), less often water. In the first case, flowers generally have a well developed and coloured perianth. In the second case, there is no perianth or it is reduced and uncoloured.
Embryo sac in Angiosperms
The diploid megaspore mother cell gives after meiosis 4 haploid megaspores from which only one survives and gives the 8 haploid cells of the embryo sac after 3 mitosis. The 8 haploid cells of the embryo sac are: – An egg surrounded by 2 synergids at the micropilar end of the ovule, – 3 antipodals at the chalazal end of the ovule, – 2 polar nuclei of the central cell at the centre of the embryo sac. These 2 polar nuclei can combine before fertilisation. 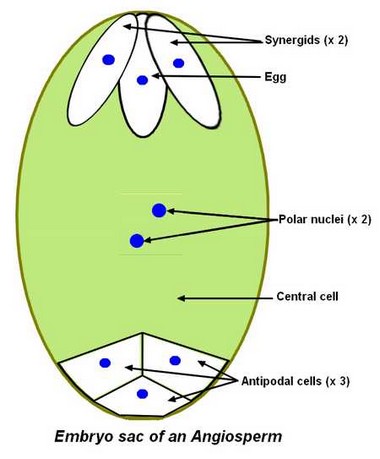
Angiosperm Ovule
Going inwards the ovule is made of: – Two integuments, the outer one being vascular. At the ovule apex, there is a tiny opening in integuments called micropyle. – The nucellus is a diploid parenchymatous tissue, surrounded by integuments and containing the embryo sac. If the nucellus is thin, the ovule is tenuinucellate; if it is thick, it is crassinucellate. – The embryo sac, the female gametophyte, is made of 8 haploids cells. 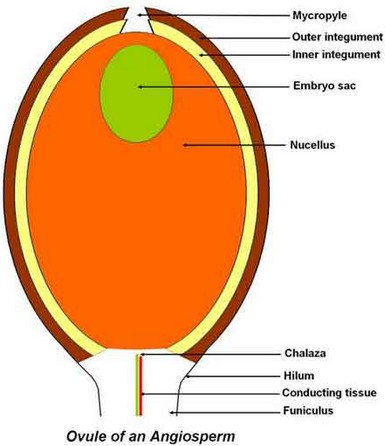
Pollen grain of Angiosperms
The pollen grain of pollen is generally made of only 2 haploid cells: – The tube cell, responsible for the elongation of the pollen tube, – The generative cell that will give 2 male gametes or sperm cells. The pollen grain is surrounded with 2 walls: – The exine, thick, ornamented or smooth, – And the intine, fine and doubling inside the exine. In certain places called apertures, exine is thinner and intine thicker. In the pollen of Monocots or Basal Dicots, there is normally only one aperture, circular or furrowed. Sometimes, this aperture stretches, being thinned by places it then gives several pores. In the pollen of true Dicots (Eudicots), there are normally 3 apertures. However, this number can vary and be multiplied. 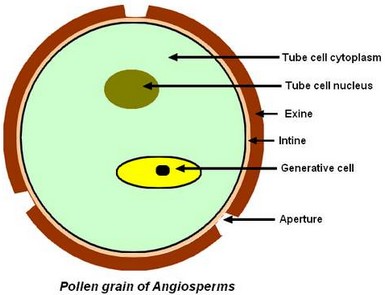
Pollen germination and pollen tube
Germination of pollen begins when the grain swells because of the absorption of the water at stigma surface. Turgescent vacuoles push the intine and the cytoplasm towards an aperture of the pollen grain of pollen: it is the beginning of the growth of the pollen tube. When the pollen tube gets longer, the 2 cells (generative cell and tube cell) and the cytoplasm are confined at its extremity, the generative cell being slightly behind the tube cell. Phenomena of cellular strain are limited at the end of the tube. The pollen tube crosses the style without ever penetrating into cells; cells of the conducting tissue act as rails for the pollen tube that finishes its growth at the level of the micropyle into the ovule.