Dry and dehiscent fruits open at maturity permitting the release of enclosed seeds. A dry and dehiscent fruit is: – A follicle when it is many-seeded, formed from one carpel and dehiscing along one side only, – A pod or a legume when it is a follicle dehiscing along two sides, – A capsule when it is formed from more than one carpel. Dry and indehiscent fruits do not split at maturity to release seeds. It is the whole fruit that is dispersed by wind or insects. A dry and indehiscent fruit is: – An achene when it is indehiscent and 1-seeded with the seed coat distinct from the fruit wall (pericarp), – A samara when it is an achene with a flat wing formed from the pericarp, – A caryopsis when it is an achene with the seed coat (testa) united to the fruit wall. It is a feature of the grasses.
All posts by _
Angiosperm carpel
A good understanding of carpel organisation is imperative to analyse transformations that occur in an Angiosperm flower after fertilisation to obtain a fruit. A carpel is made of the 3 following parts: – a stigma, a wet surface with papillate cells, that is adapted for the reception and germination of pollen (see pages on pollen), – a style, more or less developed. The pollen tube grows between its cells during the pollen germination (see pages on pollen germination), – and, an ovary, the hollow and basal part of the carpel that contains inside its cavity an ovule (see pages on ovule). Inside ovule, there is the embryo sac (see pages on embryo sac). 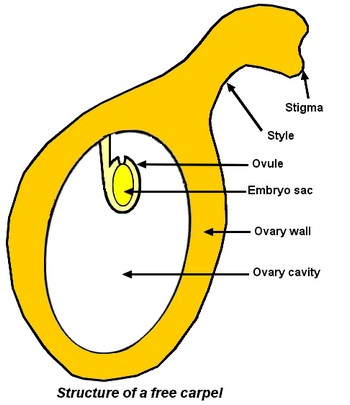 Carpels can be free and gynaecium is then apocarpous. If ovaries are entirely fused together, gynaecium is syncarpous (see pages on syncarpous gynaecium). After fertilisation of the egg contained in the embryo sac (see pages on double fertilisation), many transformations occur: – the ovary develops into the fruit and the ovary wall gives the fruit wall (pericarp), – the ovules develop into the seeds and ovule integuments give seed coat, – the fertilised egg develops in the embryo that will be in the seed. Embryo will develop in a new plant during germination.
Carpels can be free and gynaecium is then apocarpous. If ovaries are entirely fused together, gynaecium is syncarpous (see pages on syncarpous gynaecium). After fertilisation of the egg contained in the embryo sac (see pages on double fertilisation), many transformations occur: – the ovary develops into the fruit and the ovary wall gives the fruit wall (pericarp), – the ovules develop into the seeds and ovule integuments give seed coat, – the fertilised egg develops in the embryo that will be in the seed. Embryo will develop in a new plant during germination.
Double fertilisation in Angiosperms
When the pollen tube penetrates the embryo sac at the micropylar end, it goes through one synergid, burst and release the 2 sperm cells. One sperm cell and the egg fuse into a main zygote, diploid, that will give the seed embryo. The other sperm cell fuses with 2 well individualized or already fused polar nuclei. It gives a secondary zygote or endosperm that is triploid. It is called albumen in French to make the distinction with the Gymnosperms endosperm that is diploid tissue. Endosperm is usually triploid but its ploidy number can be different given that the number of polar nuclei can vary. Endosperm is a nutritive tissue which ends its development before that of the main zygote. When this last one will have ended its own development, endosperm has disappeared (seeds of Asteraceae or Fabaceae) or remains to go on its nutritive functions for the mature embryo during seed germination (seeds of Ricinus, seeds of Coco with a liquid endosperm).
Angiosperm pollination
Pollination can be made according to various modes: – Self pollination. The stigma of a flower receives the pollen of the same plant. This mode is frequent, but not compulsory, in cultivated Grasses. It is on the other hand compulsory for flowers that do not open (cleistogamous ) such as the Violet. – Crossed pollination. The stigma of a flower receives the pollen of another plant. Cross pollination can be promoted: – By dioecism: male flowers and female flowers are on separate plants (dioecious species), – By dichogamy: male and female organs mature at different times. The pollen is before released while the stigma is immature (protandry) or the stigma is receptive while stamens are still young (protogyny), – By hercogamy: some structures prevent pollen from being transferred on stigma of the same flower (rostellum of the Orchis), – By heterostyly: in Primula, flowers with high style and stamens situated on the base of the corolla must be pollinated by flowers with short style and stamens situated on the top of the corolla, – By self sterility: flowers can’t be self pollinated because of dimorphism in pollen grains and stigma surfaces. Means of pollination are the wind (anemophily) or insects (entomophily), less often water. In the first case, flowers generally have a well developed and coloured perianth. In the second case, there is no perianth or it is reduced and uncoloured.
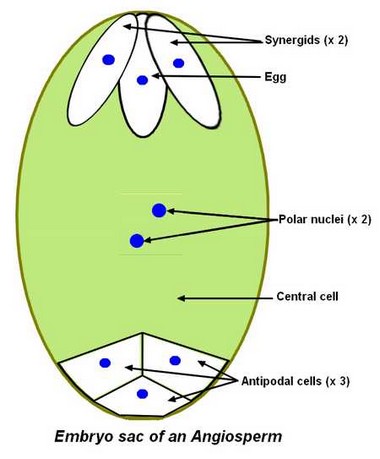
Embryo sac in Angiosperms
The diploid megaspore mother cell gives after meiosis 4 haploid megaspores from which only one survives and gives the 8 haploid cells of the embryo sac after 3 mitosis. The 8 haploid cells of the embryo sac are: – An egg surrounded by 2 synergids at the micropilar end of the ovule, – 3 antipodals at the chalazal end of the ovule, – 2 polar nuclei of the central cell at the centre of the embryo sac. These 2 polar nuclei can combine before fertilisation.